הרצאה 11-14 - הסינפסה
הסינפסה הכימית:
- מהי סינפסה:
- נקודת התקשורת בין תאי עצב, המאפשרת העברת מידע לשם קיום פעילות מוחית.
- הסינפסה מתמירה סיגנל חשמלי (פוטנציאל פעולה) לסיגנל כימי, ובחזרה לסיגנל חשמלי בתא המקבל.
- מבנה הסינפסה:
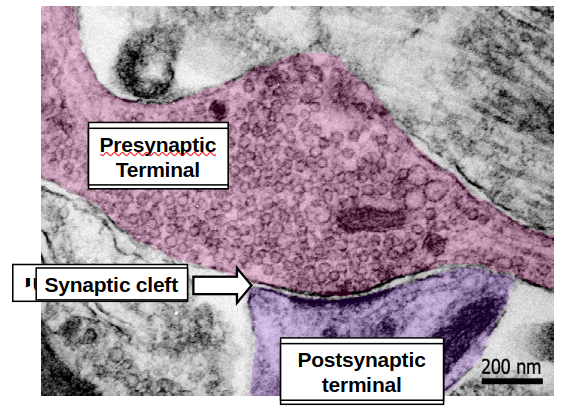
- הסינפסה היא אברון דו - תאי המורכב משלושה חלקים עיקריים:
- טרמינל פרה - סינפטי (Presynaptic Terminal):
- החלק בתא השולח, לרוב ממוקם באקסון.
- מכיל וסיקולות האוגרות בתוכן את הנוירוטרנסמיטור.
- מרווח סינפטי (synaptic cleft):
- המרווח הפיזי בין התאים אליו משוחרר החומר הכימי.
- טרמינל פוסט-סינפטי (Postsynaptic Terminal):
- לרוב נמצא על הדנדריטים.
- החלק בתא המקבל את המסר (הנוירוטרנסמיטורים).
- טרמינל פרה - סינפטי (Presynaptic Terminal):
- הסינפסה היא אברון דו - תאי המורכב משלושה חלקים עיקריים:
תהליך ההעברה הסינפטית (Neurotransmission):
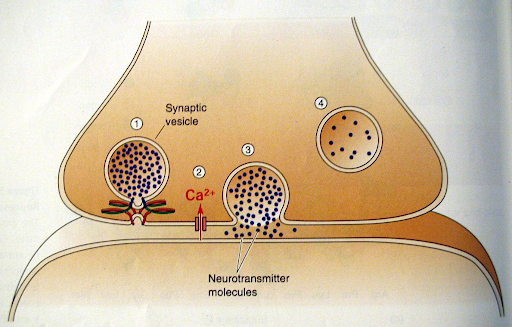
- הגעת פוטנציאל הפעולה:
- מסר חשמלי מתקדם באקסון ומגיע לטרמינל הפרה-סינפטי, וגורם בו לדפולריזציה.
- פתיחת תעלות סידן (Ca+2):
- בעקבות הדפולריזציה נפתחות תעלות סידן תלויות מתח.
- סידן נכנס פנימה אל תוך התא הפרה-סינפטי הודות למפל הריכוזים (ריכוז הסידן מחוץ לתא גדול בהרבה מזה שבפנים).
- שחרור נוירוטרנסמיטר:
- כניסת הסידן גורמת לאיחוי הוסיקולות עם ממברנת התא.
- תוחלת הוסיקולות (הנוירוטרנסמיטר) מופרשת אל המרווח הסינפטי.
- הוסיקולות עצמן אינן עוברות לתא הבא, וגם הנוירוטרנסמיטר לרוב אינו נכנס לתוכו בשלב זה.
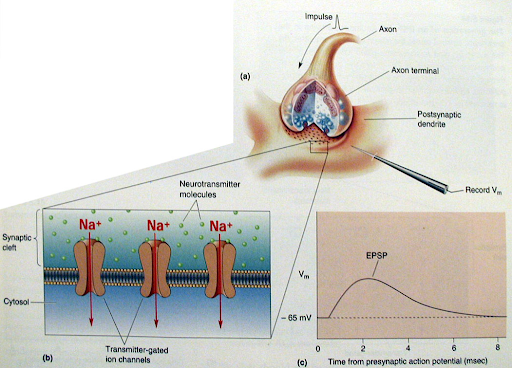
- קשירה לרצפטורים:
- הנוירוטרנסמיטור חוצה את המרווח ונקשר לרצפטורים, שהם תעלות יוניות תלויות ליגנד, הנמצאים על הממברנה הפוסט סינפטית.
- שינוי פוטנציאל הממברנה:
- פתיחת הרצפטורים מאפשרת מעבר של יונים (כמו נתרן ואשלגן) פנימה והחוצה, מה שיוצר זרם חשמלי (דפולריזציה) בתא הפוסט-סינפטי, ומתמיר את האות הכימי בחזרה לחשמל.
- הסינפסה אינה פועלת באופן בינארי כמו פוטנציאל פעולה (יורה או לא), אלא יוצרת שינוי מתח הדרגתי (גרדיאנט).
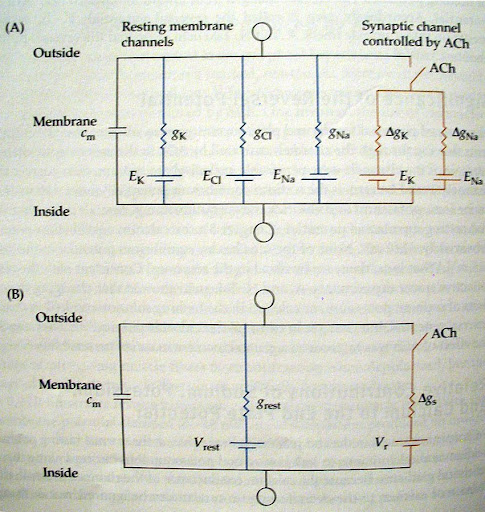
המודל החשמלי של הסינפסה ופוטנציאל היפוך:
-
מודל חשמלי והשפעה לא לינארית:
- פתיחת תעלות בסינפסה משולה להוספת נגד וסוללה למעגלי החשמלי של הממברנה.
- רצפטורים תלויי ליגנד, כמו אלו של אצטיכולין או גלוטמט, מאפשרים לרוב מעבר של קטיונים במקביל דרך אותה תעלה (למשל נתרן פנימה ואשלגן החוצה)
- ההשפעה של הסינפסה על מתח הממברנה אינה לינארית, פוטנציאל הממברנה תמיד ישאף להגיע לערך "בטריית הסינפסה" אך לעולם לא יחרוג ממנו. גם אם נפעיל את הסינפסה בעוצמה או נפעיל מספר סינפסות במקביל.
-
פוטנציאל היפוך:
- פוטנציאל ההיפוך הוא המתח (ערכה של בטריית הסינפסה) שבו הזרם נטו של היונים העוברים דרך התעלות הסינפטיות מתאפס.
- למשל: כאשר זרם הנתרן פנימה שווה בדיוק לזרם האשלגן החוצה.
- במערכות פיזיולוגיות מתח זה לרוב קרוב ל - 0mV, והוא מהווה ממוצע משוקלל של פוטנציאלי נרנסט של היונים שעוברים בתעלה.
- מדידה באמצעות קיבוע מתח (Voltage Clamp):
- ניתן למצוא את פוטנציאל ההיפוך בניסוי בו מקבעים את פוטנציאל הממברנה לערכים שונים ומפעילים את הסינפסה.
- הזרם הסינפטי הנרשם הוא לינארי ביחס למתח.
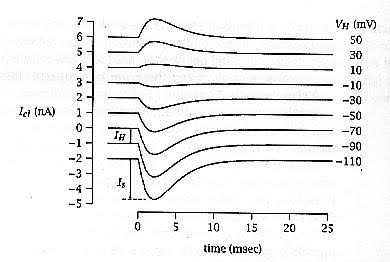
- הנקודה שבה קו הזרם הסינפטי חותך את ציר ה - X (זרם אפס) היא פוטנציאל ההיפוך הסינפטי.
- הנקודה בה זרם ההחזקה (הזרם הנדש לקבע את הממברנה ללא סינפסה) חותך את ציר ה - X היא פוטנציאל המנוחה.
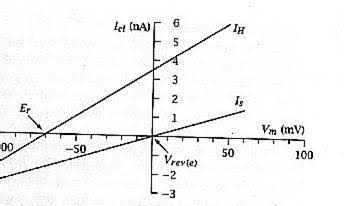
- פוטנציאל ההיפוך הוא המתח (ערכה של בטריית הסינפסה) שבו הזרם נטו של היונים העוברים דרך התעלות הסינפטיות מתאפס.
סינפסה "אמיתית" והקינטיקה של הרצפטורים:
- מודל המדרגה לעומת המציאות:
- במודל החשמלי הפשוט, הסינפסה מתנהגת כמו מתג של פונקציית מדרגה, אך בסינפסה אמיתית הקישור של הנוירוטרנסמיטר לרצפטור הוא תהליך כימי הסתברותי התלוי בריכוז החומר במרווח הסינפטי.
- בעוד שכל תעלה בודדת נפתחת ונסגרת כמקשה אחת בצורה אקראית, בסכימה המקרוסקופית מתקבלת פתיחה מתואמת של הרצפטורים (כאשר הריכוז גבוה) וסגירה הדרגתית אקספוננציאלית ככל שהנוירוטרנסמיטר מסולק. הזרם הכללי שמתקבל נראה כמו גלי אלפא.
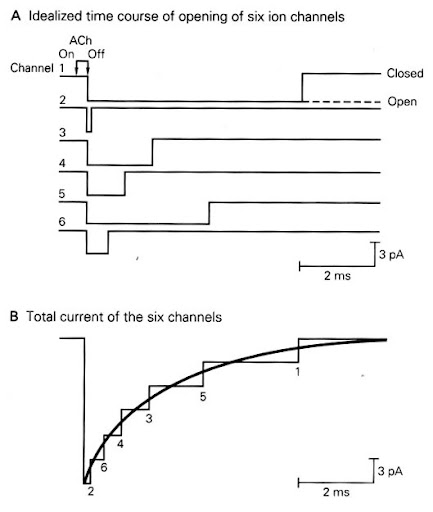
- במודל החשמלי הפשוט, הסינפסה מתנהגת כמו מתג של פונקציית מדרגה, אך בסינפסה אמיתית הקישור של הנוירוטרנסמיטר לרצפטור הוא תהליך כימי הסתברותי התלוי בריכוז החומר במרווח הסינפטי.
- סילוק הנוירוטרנסמיטר מהמרווח הסינפטי:
- סילוק מהיר של הנוירוטרנסמיטר הוא קריטי. הוא מאפשר לסינפסה להיות פעילה לזמן קצר, ובכך מאפשר לנוירונים לשדר פוטנציאלי פעולה בתדרים גבוהים (תדר המקודד למשל את עוצמת ההתכווצות של השריר) מבלי שהאותות "יימרחו".
- פירוק אנזימטי (נוירוטנסמיטור מסוג אצטילכולין):
- אצטילכולין (ACh) מפורק בצורה אגרסיבית על ידי האנזים אצטילכולין-ציטרז (AChE).
- רמת היעילות של האנזים גורמת לכך שחלק ניכר מהאצטילכולין כלל לא מספיק להגיע לצד הפוסט-סינפטי.
- קליטה מחדש (נוירוטרנסמיטור מסוג גלוטמט):
- גלוטמט מסולק על ידי טרנספורטרים של תהליך Reuptake הנמצאים בתאי הגלייה המקיפים את הסינפסה או בתאים עצמם
- בתהליך ה Reuptake הטרנספורטרים ממחזרים את הגלוטמט במקום פירוק כדי לא לגרום נזק לתא.
השפעת קבל הממברנה (ארבעת שלבי הזרם הסינפטי):
- מאחר והממברנה מתפקדת כקבל, השינוי במתח תמיד "מעוכב" ומגיע לשיאו מאוחר יותר ביחס לזרם הסינפטי הראשי.
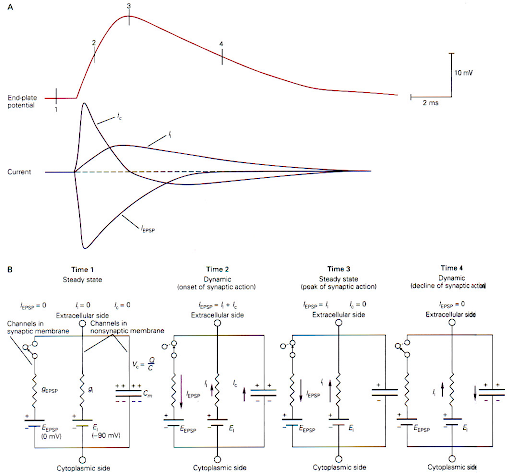
- בתמונה מעלה, ניתן לראות את חלוקת התהליך ל-4 שלבים (משמאל לימין):
- מצב מנוחה (Steady state):
- הסינפסה סגורה. הזרם הסינפטי והקיבולי אפס
- התפתחות הפוטנציאל (Dynamic onset):
- הרצפטורים נפתחים ויש זרם סינפטי גדול פנימה (של יוני נתרן).
- הזרם מתפצל לשניים:
- חלק הולך לטעינת הקבל (זרם קיבולי חיובי מקדים)
- חלק זורם החוצה דרך תעלות הדלף.
- שיא הפעולה הסינפטית:
- כשהמתח מגיע לשיאו, הנגזרת שלו שווה לאפס ולכן הזרם הקיבולי מתאפס.
- בשלב זה הזרם דרך הסינפסה זהה לחלוטין לזרם הדלף והפוך לו.
- דעיכה (Decline):
- התעלות הסינפטיות נסגרות.
- כעת הקבל פורק את מטענו לאט דרך תעלות הדלף ההתנגדותיות, ולכן הדעיכה איטית בהרבה משלב העלייה.
- מצב מנוחה (Steady state):
התפשטות חשמלית ואינטגרציה בתא שאינו איזופוטנציאלי:
- רוב הנוירונים אינם איזופוטנציאליים והמתח בהם אינו אחיד.
- השפעת המרחק החשמלי (תורת הכבל):
- אות סינפטי שנוצר בדנדריט דועך בצורה פסיבית ככל שהוא מתקדם (בשל דליפת זרם דרך הממברנה). ככל שהסינפסה רחוקה יותר חשמלית, המתח שיגיע ל-Axon Hillock (האזור בו הסומה מתחברת אל האקסון) יהיה קטן ומרוח יותר.
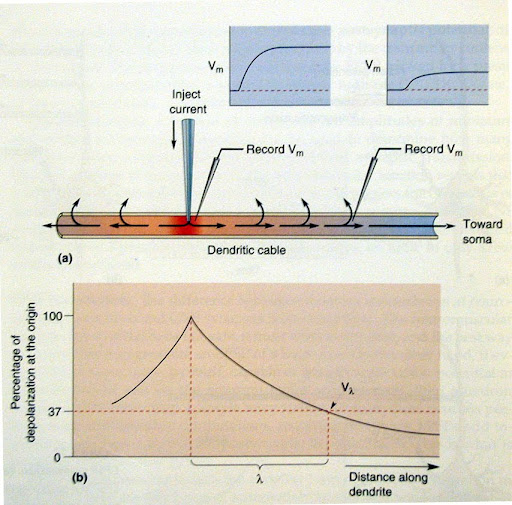
- אינטגרציה סינפטית: התא "סוכם" את המידע שמגיע אליו בשתי דרכים עיקריות:
- סכימה מרחבית (Spatial Summation):
- פוטנציאלים מסינפסות שונות הפועלות במקביל על ענפים שונים, מתחברים יחד (בצורה לא לינארית) בגוף התא.
- סכימה זמנית (Temporal Summation):
- הפעלה רציפה ומהירה של אותה סינפסה גורמת לאות החדש להיבנות על גבי פוטנציאל הממברנה שטרם דעך.
- סכימה מרחבית (Spatial Summation):
סינפסות אקסיטטוריות ואינהיביטוריות:
- הגדרות פונקציונליות:
- אקסיטטורית:
- סינפסה ש פוטנציאל ההיפוך (משוואת גולדמן) שלה נמצא מעל לערך הסף. תייצר לרוב דפולריזציה (EPSP).
- אינהיביטורית:
- סינפסה שפוטנציאל ההיפוך שלה נמצא מתחת לערך הסף. מטרתה להפריע לממברנה להגיע לסף (IPSP).
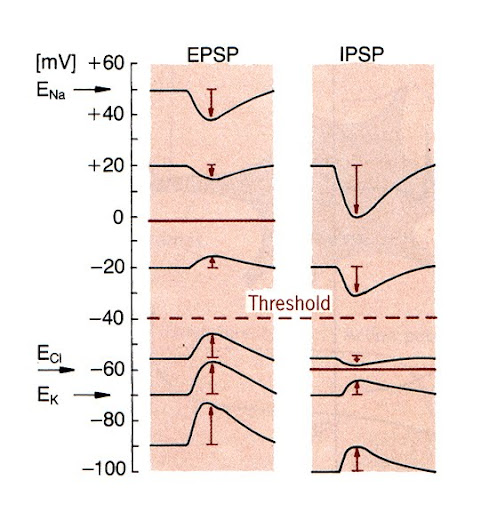
- אקסיטטורית:
- אינהיביציה מסוג Shunting (מיצד):
- סינפסות אינהיביטוריות רבות (כמו רצפטורי GABAA) פותחות תעלות כלור.
- מאחר שפוטנציאל ההיפוך של כלור קרוב לפוטנציאל המנוחה, הפעלתן בלבד לא בהכרח תשנה את מתח התא נראית לעין.
- עם זאת, פתיחתן מקטינה משמעותית את התנגדות הכניסה של התא. כאשר זרם אקסיטטורי יגיע, הוא "ידלוף" כקצר החוצה, וההשפעה האקסיטטורית תצומצם.
- השפעת מיקום ותזמון:
- מיקום:
- סינפסות אינהיביטוריות ממוקמות פעמים רבות קרוב לסומה או על הענפים הראשיים. משם הן חותכות פוטנציאלים אקסיטטוריים חלשים שמגיעים מהקצוות.
- תזמון:
- אינהיביציה המופעלת קצת אחרי האקסיטציה מקצרת את משך ה-EPSP על ידי יצירת "דלף" גדול יותר לפריקת הקבל.
- מיקום:
הבסיס הקוונטאלי של התקשורת הסינפטית:
- השערת היחידה:
- וזיקולות הן בעלות קוטר יחסית קבוע ומכילות כמות דומה של נוירוטרנסמיטרים, לכן ניתן לשער שהשחרור מבוסס על "יחידות יסוד" בלתי ניתנות לחלוקה.
- השפעת ריכוז הסידן (Ca2+) בתמיסה:
- זרם הסידן הוא הטריגר לשחרור. לכן, הורדת הסידן החוץ-תאי מקטינה את משרעת השחרור (ה-EPP).
- בריכוזי סידן נמוכים במיוחד, תגובת השחרור הופכת לכפולות שלמות של המשרעת הבסיסית (mEPP), ולעיתים יש כשלונות מוחלטים.
- מכך הוסק כי אירוע סינפטי רגיל מורכב מהמון יחידות בסיס ("קוונטות") שנשפכות יחדיו בתהליך הסתברותי.
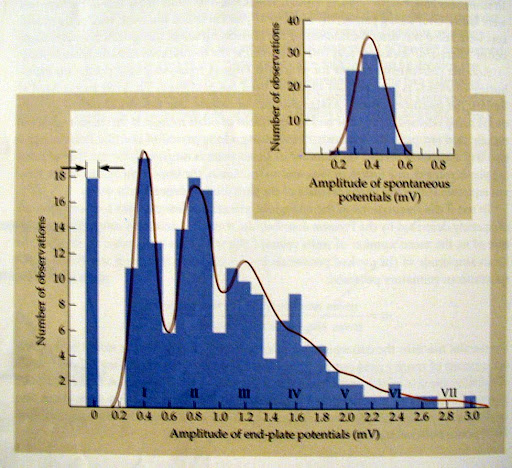
פסיליטציה קצרת מועד (Paired Pulse Facilitation) וסידן שאריתי:
- הגברת התגובה הסינפטית:
- כאשר נותנים שני גירויים לטרמינל הפרה-סינפטי בהפרש זמנים קצר, התגובה הפוסט-סינפטית לגירוי השני גדולה יותר מהתגובה לגירוי הראשון.
- תופעה זו היא סוג של פלסטיות קצרת מועד הנקראת פסיליטציה (Facilitation).
- תיאוריית הסידן השאריתי (Residual Calcium):
- העלייה בשחרור הנורוטרנסמיטר בגירוי השני נובעת מכך שסידן מהגירוי הראשון נכנס לתא וטרם סולק במלואו.
- הסידן השאריתי מצטבר יחד עם הסידן שנכנס בגירוי השני, מה שמגדיל את ההסתברות לשחרור וזיקולות.
- ככל שמרווח הזמן בין הגירויים גדל, למנגנוני הפינוי יש יותר זמן לסלק את הסידן, והאפקט של הפסיליטציה קטן.
האתר הפעיל, מנגנון איחוי הוזיקולות וקומפלקס ה-SNARE:
- האתר הפעיל (Active Zone) והכנת הוזיקולות:
- וזיקולות המוכנות לשחרור מעוגנות על הממברנה באזור הנקרא "האתר הפעיל", בסמוך מאוד לתעלות הסידן.
- תהליך ההכנה כולל את שלב הקישור הראשוני (Tethering), ואחריו את שלב העיגון והדריכה (Docking & Priming) שבו נוצר קומפלקס חלבונים יציב שאוגר את האנרגיה הדרושה לאיחוי.
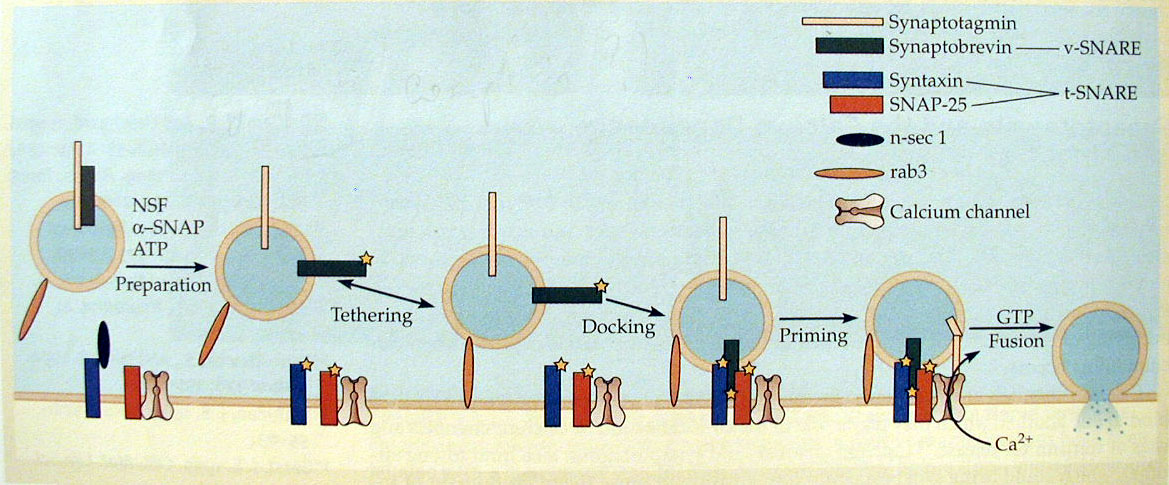
- קומפלקס ה-SNARE:
- מורכב מחלבונים היוצרים מבנה סלילי (אלפא הליקס) שמתלפף כמו קפיץ:
- חלבון ה v-SNARE: שמצוי על הוזיקולה
- חלבוני הt-SNARE: שמצויים על ממברנת המטרה.
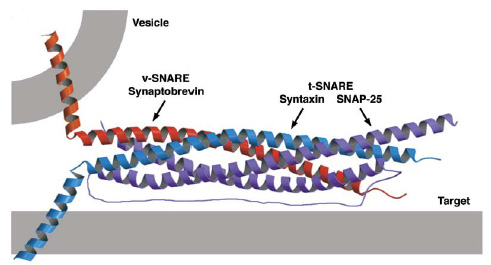
- מורכב מחלבונים היוצרים מבנה סלילי (אלפא הליקס) שמתלפף כמו קפיץ:
- איחוי ופירוק הקומפלקס:
- לפני האיחוי הקומפלקס נקרא Trans-SNARE.
- כניסת סידן מפעילה את הגלאי על הוזיקולה הנקרא סינפטוטגמין, שנותן את הפקודה לשחרר את ה"מעצור" לאיחוי הממברנות ושחרור התוכן.
- לאחר האיחוי, הקומפלקס הופך ל-Cis-SNARE (כאשר כל המרכיבים על אותה הממברנה). מכיוון שזהו קומפלקס יציב מאוד באנרגיה נמוכה, יש צורך בחלבון NSF (בעל מבנה טבעת משושה) שמשקיע אנרגיה כדי לפרק את הקומפלקס לקראת מחזור המרכיבים.
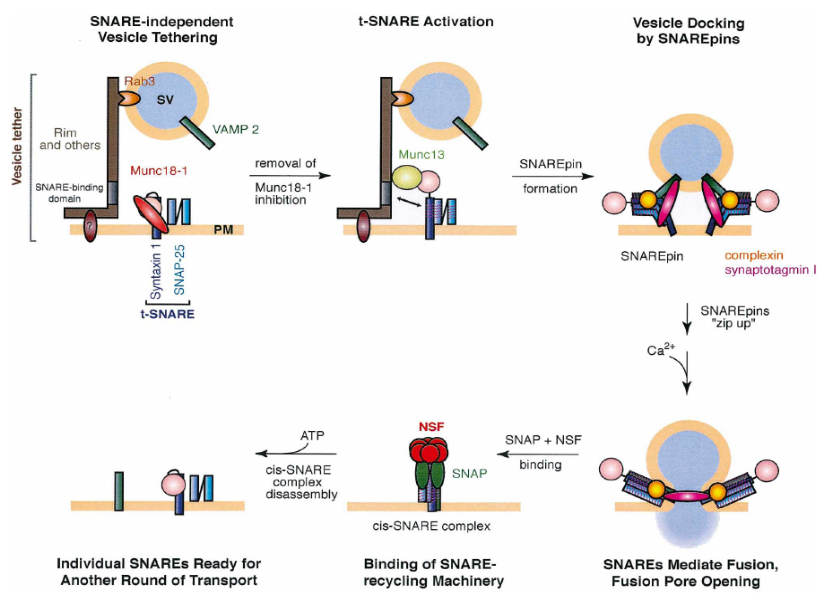
מחזור הוזיקולות (אנדוציטוזה) והטענתן מחדש
- אנדוציטוזה (יצירת וזיקולה חדשה):
- הוזיקולות החדשות נוצרות מהממברנה הפלזמטית על ידי חלבון בשם קלאטרין.
- הקלטרין יוצר מבנה קשיח ומאורגן של משושים ומחומשים (בצורת כדורגל), מה שמכתיב את הקוטר האחיד של הוזיקולות.
- אדפטורים (מתאמים) המקשרים בין הממברנה לקלאטרין בוררים אילו חלבונים ספציפיים (כגון v-SNARE) ייכנסו לוזיקולה החדשה ואילו יישארו בחוץ. לבסוף, הוזיקולה ניתקת מהממברנה והכלוב של הקלאטרין מפורק.
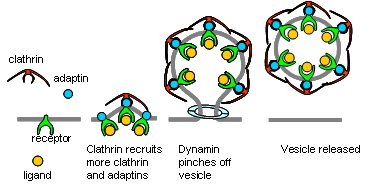
- הטענת הנורוטרנסמיטר (שלב כפול):
- משאבת פרוטונים:
- מפרקת ATP על מנת לשאוב פרוטונים (H+) לתוך הוזיקולה, מה שיוצר סביבה פנימית חומצית (pH נמוך) ומפל ריכוזים.
- משחלף וזיקולרי (Vesicular Transporter):
- מנצל את האנרגיה הפוטנציאלית של מפל הפרוטונים (יציאתם החוצה) כדי לשאוב נורוטרנסמיטרים (כגון גלוטמט או GABA) פנימה לתוך הוזיקולה.
- המשחלף הוזיקולרי הוא החלבון המגדיר בפועל את סוג התא (למשל, תא גאבא-ארגי או גלוטמט-ארגי), שכן הוא זה שקובע איזה סוג של נורוטרנסמיטר יאוכסן בוזיקולות ויופרש.
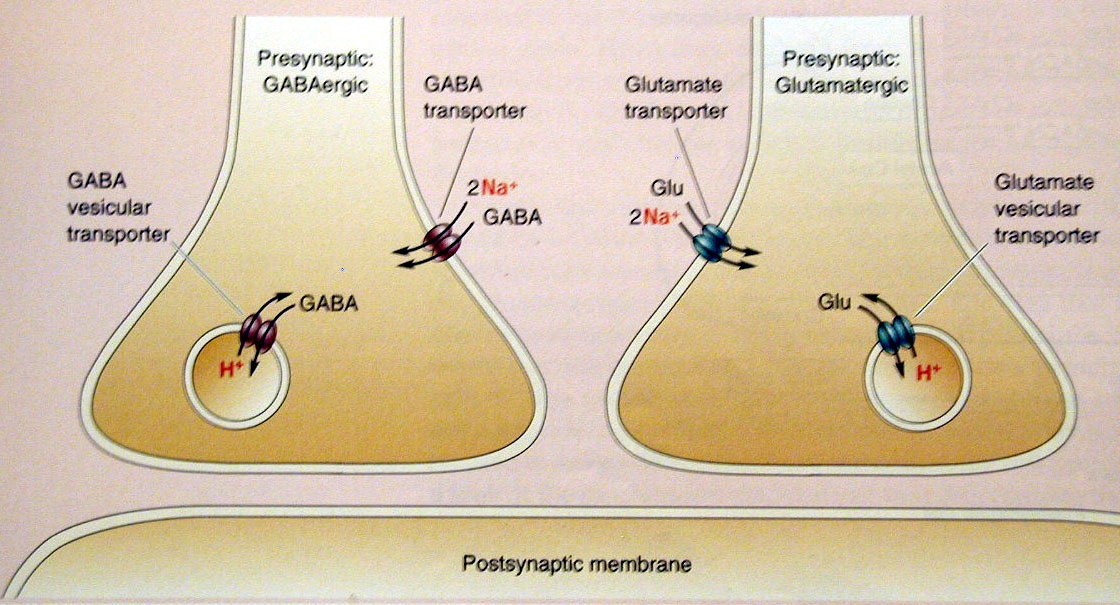
- משאבת פרוטונים:
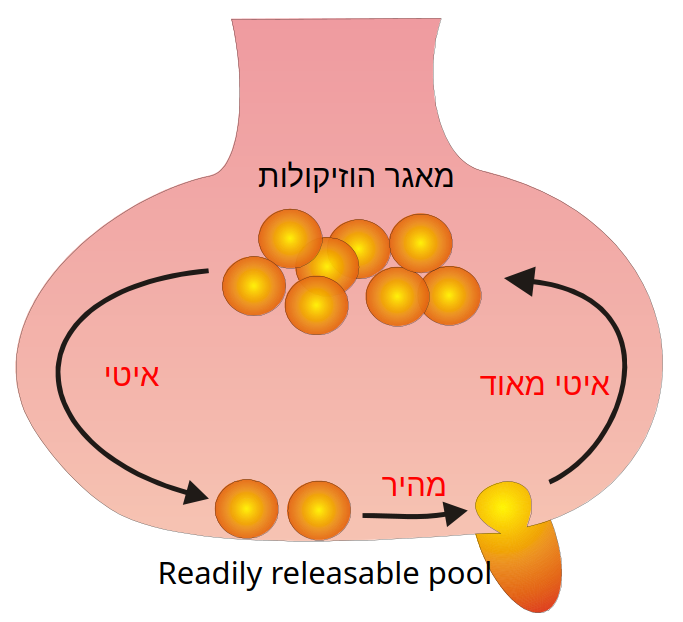
מאגרי הוזיקולות ודיכוי סינפטי קצר מועד (Short Term Depression)
- מאגרי הוזיקולות בטרמינל:
- אזור ה- Readily Releasable Pool (RRP): וזיקולות שמוכנות לשחרור מיידי ומעוגנות באתר הפעיל.
- אזור ה - Reserve Pool: מאגר רזרבי גדול הנמצא מאחור.
- דיכוי קצר מועד (Short Term Depression):
- כאשר הטרמינל מגורה בתדר גבוה ורציף, התגובה הפוסט-סינפטית הולכת וקטנה. הסיבה לכך היא שהשחרור של הוזיקולות מה-RRP הוא תהליך מהיר, אך הגיוס והמחזור שלהן בחזרה (מהרזרבה ומאנדוציטוזה) הוא תהליך איטי יותר.
- לכן, הגירוי התדיר מרוקן את מאגר הוזיקולות הזמינות בטרם הן מספיקות להתמלא מחדש. הפסקה של הגירוי תאפשר למאגרים להתמלא מחדש והסינפסה תתאושש (מכאן שזהו דיכוי קצר מועד).
- הקשר לפרמטרים הקוונטליים (
לעומת ): - בניגוד לפסיליטציה, בה יש עלייה בשחרור הנגרמת מעלייה ב-
(הסתברות השחרור עקב סידן שאריתי), בדיכוי (Depression) השינוי הדומיננטי הוא ירידה דרסטית ב- (מספר הוזיקולות הזמינות באתר השחרור שהולכות ונגמרות).
- בניגוד לפסיליטציה, בה יש עלייה בשחרור הנגרמת מעלייה ב-
פוטנציאציה ארוכת טווח - Long Term Potentiation:
- שינוי יציב וארוך טווח בחוזק הסינפטי (הגברת התגובה הפוסט-סינפטית), המהווה את הבסיס התאי לתהליכי למידה וזיכרון.
- מאפיינים:
- ספציפיות (Specificity):
- ה - LTP אינו פועל על כל העץ הדנדריטי, אלא מחזק רק את הסינפסות הספציפיות שהיו פעילות באותו הזמן.
- סינפסות שכנות על אותו דנדריט שלא שוחרר בהן נוירוטרנסמיטור, לא יתחזקו.
- אסוציאטיביות (Associativity):
- תנאי הכרחי ל - LTP הוא שילוב של שחרור נוירוטרנסמיטר מהצד הפרה-סינפטי, יחד עם דפולריזציה חזקה של התא הפוסט-סינפטי.
- סינפסות שפעילות בו זמנית עם יריית התא הפוסט-סינפטי יתחזקו (חוק הב - Hebb's rule).
- ספציפיות (Specificity):
הרצפטורים היונוטרופיים לגלוטמט:
- ההבנה של מנגנון ה - LTP נשענת על ההבדלים בין שתי קבוצות רצפטורים אקסיטטורים המגיבים לגלוטמט.
- רצפטורי AMPA:
- תעלות מהירות ופשוטות.
- המוליכות שלהן אינה תלויה במתח הממברנה, כל עוד גלוטמט נקשר אליהן, התעלה נפתחת.
- רצפטורי NMDA:
- נפתחים ונסגרים לאט יותר
- הם גם תלויי ליגנד וגם תלויי מתח. במתח המנוחה של התא, פתח התעלה חסום פיזית על ידי יון מגנזיום.
- כדי לאפשר זרם, נדרש גם שגלוטמט ייקשר וגם שהממברנה תעבור דפולריזציה חזקה מספיק כדי להוציא את יון המגנזים החוצה.
- חדירות ליונים:
- בעוד ש - AMPA מעביר בעיקר נתרן ואשלגן, NMDA מאפשר גם כניסה של יוני סידן.
מנגנון האינדוקציה של Early LTP:
- כאשר התא הפוסט-סינפטי חווה דפולריזציה וגלוטמט משתחרר למרווח, תעלות NMDA נפתחות ומכניסות סידן.
- הסידן משמש כשליח משני, הוא נקשר לחלבון הקלמודולין ויחדיו הם מפעילים קינאז ספציפי בשם CaMKII. קינאז זה מזרחן את עצמו וכך נשאר פעיל לזמן מה.
- הגברת התגובה הסינפטית:
- הקינאז CaMKII מזרחן את רצפטורי ה - AMPA הקיימים ובכך מגדיל את מוליכותם.
- מעבר לכך, במקום שרצפטורי ה - AMPA יעברו אנדוציטוזה טבעית, הזרחון מגן עליהם ומוביל להחדרה והצגה של יותר רצפטורי AMPA לממברנה באזור הסינפסה.
- ככל שיש יותר AMPA, התא יגיב בעוצמה חזקה יותר לאותה כמות של גלוטמט.
- סינפסות שקטות:
- קיימות בממברנה סינפסות המכילות רק רצפטורי NMDA ללא רצפטורי AMPA.
- במנוחה, סינפסות אלו לא מגיבות לגלוטמט ולכן "אינן קיימות פונקציונלית".
- רק בתהליך ה - LTP, מוחדרים אליהן רצפטורי AMPA והן הופכות לפעילות לראשונה
- פוטנציאל פעולה חוזר (Back-propagating Action Potential):
- כדי שסינפסות רחוקות יחוו דפולריזציה חזקה מספיק לשחרור חסימת ה - NMDA, הן מסתמכות על התפשטות אחורנית של פוטנציאל הפעולה.
- כאשר התא יורה פוטנציאל פעולה מאזור ה - Axon Hillock, הדפולריזציה אינה עוברת רק לאקסון, אלא חוזרת באופן אקטיבי חזרה לתוך העץ הדנדריטי, המכיל אף הוא תעלות נתרן וסידן תלויות מתח.
- עקרון האסוציאטיביות:
- תופעת פוטנציאל הפעולה החוזר קריטית לעקרון זה.
- העקרון הוא "דיווח" לכל הסינפסות בעץ שהתא ירה פוטנציאל פעולה הרגע. סינפסה שתקלוט גלוטמט באותו שבריר שנייה בדיוק, תתחזק.
מידור אזורי - עוקצים דנדריטיים (Spines):
- רוב הסינפסות האקסיטטוריות נוצרות על בליטות קטנות הנקראות עוקצים דנדריטיים, אשר מרחיקים את נקודת הקשר מהגזע המרכזי של הדנדריט.
- מחסום דיפוזיה:
- הצוואר הצר של העוקץ מונע מהסידן הנכנס להתפזר בחופשיות לענפי הדנדריט ולסינפסות השכנות.
- מידור זה מבטיח שהפעלת CaMKII תתרחש מקומית בלבד וכך מבטיחה את ספציפיות המערכת.
- העוקץ מאפשר הולכה חשמלית (דפולריזציה חודרת פנימה), אך מגביל מעבר של יונים וחלבונים.
קונסולידציה (Late LTP) ויצירת זיכרון ארוך טווח:
- ה- LTP המוקדם הוא הפיך ודועך לרוב לאחר כמה שעות.
- על מנת "למצק" את החוזק הסינפטי לכדי LTP ארוך טווח (קונסולידציה), נדרשת סינתזת חלבונים חדשים ושעתוק mRNA.
- תרגום חלבון מקומי בדנדריטים:
- המוח מייעל את העברת החלבונים על ידי יצירת mRNA בגוף התא ואריזתו בגרנולות (RNA Granules), המגנות עליו מפני פירוק, אותן הוא משנע לאורך העץ הדנדריטי לאזור הסינפסה הפעילה.
- כאשר ה - RNA מגיע לאזור סינפסה שעברה LTP, ריבוזומים מקומיים מתרגמים אותו מידית לחלבונים, אשר תורמים להרחבה פיזית משמעותית בגודל העוקץ וליצירת קשרים סינפטיים חדשים.
דיכוי סינפטי ארוך טווח (LTD - Long Term Depression):
- משמש כדי לאזן את הרשת ולמנוע מצב של היפר - אקסיטביליות (תופעה שעלולה לגרום לאפילפסיה).
- מחליש באופן ספציפי סינפסות שפעילותן חלשה ושומר על משקלים יחסיים בין סינפסות שונות.
- אינדוקציה מנוגדת ל- LTP:
- ה - LTD נגרם על ידי מתן גירוי ממושך בתדר נמוך לזמן ממושך.
- ההבדל בין יצירת LTP ל - LTD מבוסס על כמות הסידן הנכנסת לתא.
- ריכוז סידן גבוה:
- מפעיל פרוטאין קינאזות שמזרחנות חלבונים ומובילות ל - LTP (כניסה חזקה דרך הרבה רצפטורי NDMA).
- ריכוז סידן נמוך ורציף:
- ה - LTD מתקבל בגירוי חלש ומתמשך, ומפעיל אנזימים מסוג פוספטזות (המסירות פוספט מהחלבונים), שגורמות להסרת רצפטורים ולהחלשת הסינפסה.
- ריכוז סידן גבוה: